Technologies to map the conformation of the human genome
Topological interactions between distant DNA segments play fundamental roles in the function of eukaryotic genomes, including activation of promoters by distant enhancers, the establishment of insulated gene expression domains, and control of essential genetic recombination events. Key methodological advances have led to tremendous progress in understanding the molecular mechanisms shaping intramolecular chromosome loop structures. In contrast, while topological interactions across separate chromosomal DNA molecules are well known to contribute to essential biological processes like homology-directed repair and chromosome segregation, limitations in our ability to distinguish specific chromosome conformations across paired molecules have hindered research into the underlying mechanisms. With our recent development of sister chromatid-sensitive Hi-C (Mitter et al., Nature, 2020), which allows inter-molecular contacts between sister chromatids to be distinguished from intra-molecular contacts forming loops on individual sister chromatids, we have opened new possibilities to study topological interactions. We are currently developing next-generation scsHi-C technology and a computational framework to generate high-resolution genome-wide maps of replicated human chromosomes. We are further developing a super-resolution imaging approach to trace sister-chromatid fibers with nanometer precision, thereby assessing the conformational variability in individual cells.

Molecular mechanisms shaping human chromosomes
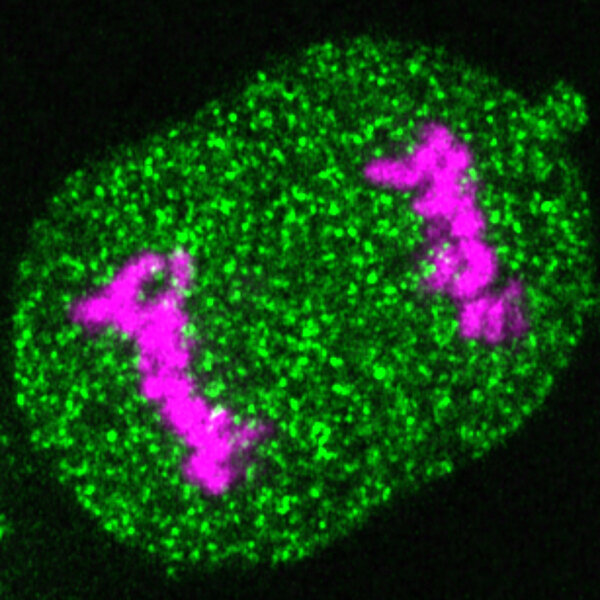
To understand how intra- and inter-molecular DNA interactions contribute to gene expression, chromosome segregation, and homology-directed recombination, we need to consider how the dynamic organization of replicated chromosomes can be generated by a complex interplay between multiple molecular activities. Within chromatin fibers, cohesin continuously forms loops whereas long-range affinity interactions between chromatin regions containing characteristic histone-modification patterns establish large-scale nuclear compartments, putatively involving microphase separation. Following DNA replication, topological linkages between sister chromatids are formed by a distinct pool of cohesin that remains stably bound to DNA. How cohesive cohesin is positioned on the genome, how it interacts with loop-extruding cohesin, and how sister-chromatid interactions are affected by chromatin-mediated compartmentalization, are poorly understood. To address these fundamental questions, we combine our sequencing- and imaging-based chromosome conformation capture technologies with precise perturbations and cutting-edge computational analysis. With this approach, we aim to integrate our detailed structural and functional data into biophysical models toward a mechanistic understanding of how chromosomes self-organize based on the complex interplay between motor-driven DNA loop extrusion, topological linkages, and chromatin microphase separation.
Physiological roles of chromosome conformation
During cell division, the highly intermingled sister chromatids are disentangled and folded into separate compact bodies to allow spindle-based movement of one genome copy to each nascent daughter cell. We have recently found that mitotic chromosomes assemble by coordinated action of condensin-mediated DNA looping and an acetylation-regulated chromatin phase transition, which confers mechanical resistance to spindle microtubules (Schneider et al., Nature, 2022). The set of mitotic chromosomes is then kept as separate bodies by a repulsive surface formed by the surfactant-like protein Ki-67 (Cuylen et al., Nature 2016). At the end of mitosis, the inactivation of Ki-67 induces the formation of a single cluster from the entire set of chromosomes to displace cytoplasm and build a single nucleus (Samwer et al., Cell, 2017; Cuylen-Haering et al., Nature, 2020). In our ongoing and future work, we aim to understand how the interplay between cohesin- or condensin-mediated DNA looping, cohesive linkages between sister chromatids, and chromatin phase separation cooperatively contribute to various processes inside the cell nucleus.
Michael Mitter: Unraveling the genome in 3D-space
For more details see: Mitter et.al., Nature, 2020. doi: 10.1038/s41586-020-2744-4
Matthias Samwer: How a set of mitotic chromosomes forms a single nucleus during mitotic exit
For more details see: Samwer et.al., Cell. doi: 170(5):956-972.e23
Sara Cuylen: How a surfactant-like protein keeps chromosomes apart during mitosis
For more details see: Cuylen et.al., Nature. doi: 535(7611):308-12
Beata Mierzwa: How ESCRT-III forms a dynamic filament system to cut cells apart during cytokinesis
For more details see: Mierzwa et.al., Nat Cell Biol. doi: 10.1038/ncb3559. Epub 2017 Jun 12.
Ana David: An amplified spindle - how molecular self-organization shapes the cell division machinery
For more details see: David et.al., Journal of Cell Biology, May 2019, doi: 10.1083/jcb.201805044