Research
Epigenetic Changes in Plants
The characteristics of organisms are influenced by two components of inheritance: genetic and epigenetic information. While the DNA sequence of the genome provides the blueprint that is transmitted from one generation to the next, epigenetic factors determine the three-dimensional organization of the genome, transcriptional activity regulating differentiation and development, and stability and defense against intruding DNA and RNA molecules. Our group is interested in the mutual influence of genetic and epigenetic determinants. We study chromatin components of DNA repair and epigenetic information under abiotic stress, during differentiation, and in polyploids, using Arabidopsis thaliana as model organism. With Aethionema arabicum, we study adaptive traits in seed formation and germination.
Response of chromatin to heat stress
Most genomes contain large repetitive regions that are poorly transcribed. These are densely packed, visible by strong accumulation of DNA stain and therefore called heterochromatin, in contrast to less stained euchromatin that contains most of the active genes. During developmental transitions and in response to different environmental conditions, heterochromatin can decondense, allowing its content to contribute to the RNA pool of cells. Among transcripts from heterochromatin are many derived from transposons, opening a chance for amplification of these “jumping genes”. We analyze changes in chromatin composition in response to heat stress, resulting in transient activation of heterochromatic genes, changes in the association of the DNA with histones, and in nuclear shape and organization (Fig. 1).

Live imaging of nuclei in the root of young Arabidopsis seedlings expressing H2B-RFP, before (T0) and during heat stress (10/20/30 h) and after recovery at regular temperature (2/7 days later). Size bar 10 μm.
Photos: Tao Dumur
Role of chromatin and RNA during repair of DNA damage
While epigenetic changes are largely reversible, mutations at the level of the DNA sequence are often permanent and mostly deleterious. Plants have several efficient repair mechanisms to deal with DNA damage. Lesions embedded in chromatin need to be made accessible to the repair enzymes, which is achieved by chromatin remodeling complexes (Fig. 2). We investigate the role of one such complex during repair of random or induced and targeted DNA damage, the interplay between the protein components as well as the role of specific RNAs.
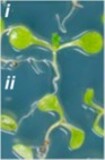

Fig. 2: Lack of chromatin remodeling impairs growth after DNA damage.
Repair-efficient young Arabidopsis seedlings can form true leaves (i) after exposure to DNA double strand break-inducing bleomyin, while repair-deficient plants cannot (ii, left). Mutants lacking components of the SWR1-C complex (pie1, arp6, swc6) are significantly impaired in their DNA damage repair, comparable to the repair-deficient ku70 mutant (right).
Images: Rosa et al. 2013
Heritability of epigenetic changes
Although epigenetic changes are often transient and reversible, there are several examples for the formation of epialleles that are stably inherited in either silent or active state even between generations, without being encoded in the DNA sequence. How these epialleles originate and what determines their stability is not clear. We investigate the nature of a pair of epialleles and their paramutation-like interaction, a case of non-Mendelian inheritance. We also focus on the epigenetic configuration in the stem cells of the shoot (Fig. 3), as potentially heritable epigenetic changes must pass through them to the progeny.

Expression of H2B-mCherry under control of the CLV3 promoter in a young Arabidopsis seedling. Whole-mount immunostaining with mCherry antibodies and laser scanning microscopy. Size bar 10 μm.
Photos: Ruben Gutzat
Light inhibition of seed germination
Timing of seed germination is crucial for seed plants and coordinated by internal and external cues, reflecting adaptations to different habitats. Light is an important factor. In contrast to Arabidopsis that requires light for efficient germination, some accessions of distantly related Aethionema arabicum germinate only in complete darkness (Fig. 4). We investigate physiology and molecular biology and a potential epigenetic basis of this light inhibition and ask whether this is an adaptive trait.

Fig. 4: Seed germination in response to light in Aethionemaarabicum.
The accession from Turkey (top) germinates equally well in light and darkness; the accession from Cyprus (bottom) is inhibited by light. Size bars 1 cm or 2 mm, respectively.
Photos: Zsuzsanna Mérai